����ժҪ��ժҪ������(oxalicacid)��һ����Ҫ�������л����㷺�ֲ���ֲ�����������У��ڲ�ͬ���������з�����Ҫ���ܡ����Ļع��˹��������������������о���չ����������ܹ����ڲ��ᣬ����ֲ�ﲡԭ�����ʳҩ���������ҵ����ȡ�������Ϊһ�ּĶ�Ԫ���ᣬ��
����ժҪ������(oxalicacid)��һ����Ҫ�������л����㷺�ֲ���ֲ�����������У��ڲ�ͬ���������з�����Ҫ���ܡ����Ļع��˹��������������������о���չ����������ܹ����ڲ��ᣬ����ֲ�ﲡԭ�����ʳҩ���������ҵ����ȡ�������Ϊһ�ּĶ�Ԫ���ᣬ���������Ҫͨ��������ѭ��;������ȩ��ѭ��;���Ͳ�������;���ϳɡ����������������һ����Ҫ���������ӣ��ܹ�Ӱ������������뷢�������ܹ�ͨ���������á��ữ������֯����������ϸ���ڽ�����յ����������������÷�ʽ������������ֲ��Ļ����з������ã������������ʷ����Ⱦѭ���з�����Ҫ������ѧ�Ͳ���ѧ���á����ĴӲ�����������ԡ���л;�����������������ࡢ��������á�������ػ����ܵȷ����������������δ����������о�Ӧ��ע����������˽��顣
�����ؼ��ʣ���������ᣬ��л���²��ԣ�в��
��������(oxalicacid��OA)��һ�ֹ㷺�����ڶ��ֲ���������������ڵ���Ҫ��л����[1]������ܹ��ϳɲ����ڲ�����ά�����滷�������˵�pHֵ[2]�������������������������(����ͷ���������)в����Ӧ�ȷ��淢����Ҫ����[3-5]��
�������﷽����ְ֪ʶ��������ת���������ķ����ڿ�
����1������������
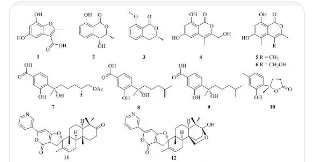
���������ֳ��Ҷ��ᣬ��Ϊ�ڶ��л�ǿ����һ����Ķ�Ԫ���ᣬ�������Ҵ���������ˮ���������ѣ������ڱ����ȷµ��л��ܼ�����������ɫ��������ĩ��ͨ���Զ�ˮ������(H2C2O42H2O)��̬��������Ȼ�磬����2�־���ṹ��̬(���ε�α�ͺ͵�б���ε�β��)������ܶ�ԼΪ1.65������������Ϊ1.54[6]��������190°C����Ũ����ʱ�ֽ�ɶ�����̼(CO2)��һ����̼(CO)��ˮ[7]���������ȱȴ���ǿԼһ���������ͨ�ԣ�Ҳ��һ��ǿ��ԭ�������ỹ���ж��Ժ�ʴ�ԣ�Ҳ��һ��ǿ���ϼ����ܹ����������(�������Ca2+)�������γɲ�����(�����Ƶ�)[2]�����У�����в���ͨ���Բ����ε���̬����[8]��
����2�����л;��
������������������һ����Ҫ�Ĵ�л����[1]������ֲ�����ϳ�;����Ҫ������ȩ��/�Ҵ���;��������Ѫ��;���Ͳ�������;��[9-10]����ͬ��ֲ��������ϳ���Ҫ��ͨ��������ѭ��(tricarboxylicacidcycle��TCA)����ȩ��ѭ��(glyoxylicacidcycle��GAC)�Ͳ�������;�� [2,11]�����в�������;����Դ��δ��ȷ[12]���������ϳɵ�����о���չ���磬1877���״α�����27������������м�����[13]��1900������ƽ�ĸ(Saccharomycescerevisiae)��Һ����������Ҳ���ֲ��ᣬ���½�������ڶ�����ܹ��ϳɲ���[14]�������Ӱ���������ϳɵ�������Ҫ����̼Դ����Դ��Ӫ�������Լ�pHֵ�Ȼ�������[11]��
�������磬�������Գ��(Cryphonectriaparasitica)����̼���Ϊ̼Դ���������в����IJ�����࣬��pHֵ�ﵽ5.0������ʱ���۴�������[15]��ͬʱ������IJ���Ҳ�����ø�йء����磬���̾�(Sclerotiniasclerotiorum)�IJ���ϳ���Ҫͨ��������������ˮ��ø(oxaloacetateacetylhydrolase��OAH)��ˮ���������(oxaloaceticacid)���ɲ���[16]�����⣬���̾��Ĺ�������ø��ϵ�ڵ��������ת��ø(carnitineacetyltransferase��CAT)Ҳ������ز���ĺϳ�[4]��
��������Ľ�����Ҫ����������������;�������У�����;���ɲ�������ø(oxalateoxidase��OXO)����ʹ����ת��Ϊ������̼(CO2)��������(H2O2)����Ҫ�����ڲ�������絭�S��˨��(Trametesochracea)��С���Ⱥڷ۾�(Tilletiacontraversa)��[17];������;����Ҫ�����ڲ�����������ù(Aspergillusniger)���ฯ�Կ�(Postiaplacenta)�ڣ�ͨ����������ø(oxalatedecarboxylase��ODC)ʹ����ת��Ϊ����(HCOOH)�Ͷ�����̼(CO2)[18]��
�������磬�ڻ���pH�����������£�����ù(.niger��pH1.1)��ODC���Ա��յ����ӣ��Խ���OA���������������������[19];�ฯ�Կ�(Postiaplacenta)ͨ��ODC������ᣬ����ϸ���ں;�˿��Χ�IJ���Ũ��[11]�����⣬������ή��;����Ϊ�б���������������ķ��Ρ�
����ͨ������ֲ���OXO��ODC����ֽ���ᣬ�����������ã�������ԭ�������Ⱦ�����ֲ���[20-21]�����磬������OXO�����ת�����ѶԲ�����нϸߵ������ԣ��������Ļ���������ǿ���Ի�ù���;��˲��Ŀ������[22]��ת�����Ҳ�������ԵĿ����˲���״[23-24]�����⣬������ODC�����ת�����Ѻ�ݫ�ĶԺ��̾����������Ŀ���[25-26]��
����3���������
����������(1877−2021��)ͳ�ƽ����������������ܹ��������ᡣ�����������������Ҫ�������Ҿ��ź͵��Ӿ��š����У�ֲ�ﲡԭ�����Ҫ�������������ס�������������ƻ���ȸ��ò�[27]����ù��(Aspergillus)��������·��ѡ����ࡢ�̲ݺͶ��ƵȻ�ù��[28]����������(Botrytis)�����������տ����Ͳˡ����������P���������������˲�[29]�ĺ��̾���(Sclerotinia)��������⣬һЩ���������������(Paxillus)[30]�����ֽ���ľ���صĹ�ҵ�����˨����(Trametes)[11]������ʳ�þ����Ǯ����(Collybia)[31]Ҳ�ܹ��������ᡣ
����4������������
����4.1Ӱ������������뷢��
�����о������������ܹ�Ӱ������������뷢��[62]��������̾�ͨ������������Ӫ�����˾��˷��������Ի�����ʵ�ֶԲ��������Ŀ�����[63-64]�����̾���������Һ�еIJ���ɴٽ�������ڵ�ϸ���ڽ���ø�Ļ��Խ������ֲ��ϸ���ڣ���Ϊ���˷����ṩӪ����Դ[65]��
�������̾����ڵIJ��ή�ͻ���pHֵ���������˿��ԭ�����ø(mitogen-activatedproteinkinase��MAPK)�Ļ��ԣ����ؾ��˵ķ���[66];�������ת��ø(carnitineacetyltransferase��CAT)��ȱʧ������̾�Ssoah1����ı��オ�ͣ�����������ڼ��٣����¾��˷���ȱ�ݡ����Ű����μ���Ⱦ��(infectioncushion)����ɥʧ[4]��ͨ���������յ���ͬԴ����ķ�����õĺ��̾�oah����ͻ������꣬����Ϊ����ϳɼ������γ���������[67-68]��
�������⣬�ฯ�Կ�(.placenta)ͨ������OXO������ᣬ��ά��Ӫ�����˿��Χ��pHֵ���������Ũ�ȹ��߶����ƾ�˿����[11]�����ỹ�ܹ������ؼ������С�ܿ�ù(Coniothyriumminitans)�ľ�˿�����������ȷ���������Ժ��̾����ؼ�������[69]��
����4.2���벡ԭ������²��Լ������ֲ�ﻥ��
��������������Ӫ����(necrotrophic)ֲ�ﲡԭ�����Ⱦֲ��Ĺؼ�����֮һ����һ����Ҫ���²���������[21,36]������ͨ���������á��ữ����ֲ����֯����������ϸ���ڽ�����յ�����������(reactiveoxygenspecies��ROS)�ȷ�ʽ������������ֲ��Ļ����з�����Ҫ����[2,51]��
����(1)�������Բ�����ж������ã��յ�����ֲ���γ��벡ԭ���(����̾�)��Ⱦ���Ƶ�֢״���������Ⱦ�����з�����Ҫ����[36]�����磬���̾�����Ⱦ���ڷ��ڴ������ᣬ�ڼ�����֯�в��ϻ���;������Ⱦ��λ����Ũ�ȵIJ������ߣ��ڰ������ֱ�Ӳ����������ã��������ϸ������[70-71]��Ӱ�컼��ֲ���澳��Ӧ����[72]�����⣬һЩֲ��(��������ס������ס�����С���)����OXO���ԣ����Խ�������ڵIJ��������ֽ�ΪH2O2���Ƚ��Ͳ���Ķ������ã�ͬʱ������H2O2Ҳ�����ڲ�ԭ��������뼰��չ[73]��
����(2)�ữ�������ͨ�����ڲ���Ӫ����������Ⱦ�����Ի���[70]�����磬���̾�����Ⱦ���ڷ��ڲ��ᣬ�ữ����ϸ�����������ڲ�ԭ����Ⱦ[74-75]�����ż���ֲ��ϸ������������ӣ�ϸ��Ĥ��֬�ʹ�����ˮƽҲ��֮���ߣ������ı�ϸ����ͨ�ԣ�����ϸ��ɥʧˮ�ֲ�����ϸ�����ṹ���ˣ�����ϸ������[16,76]��
��������ɵ���ֲ�ﱣ��ϸ��(guardcell)�Ŀ��š���ԭ������ڲ����۲��ᣬ��������(��K+)ͨ��������������(abscisicacid)�յ������رգ����ڲ�ԭ����������ֲ��[74]���ڲ�ԭ����Ⱦ�����У�������ڵIJ����յ�����ֲ���ROS��Ӧ��ϸ������������(programmedcelldeath��PCD)[21]�����⣬���������������(PO43−)��������������ԣ��������(mycorrhiza)���ڵIJ����ܹ���ǿ��������Ԫ���ܽ������[77]��
�������磬������ù(.oxalicum)�ͺ���ù(.niger)�ܹ�ͨ����������Ӫ�����Ի����������������������ܽ�[44]������������ͨ�����ڲ���������ӽ���γɲ����(ammoniumoxalate)��ʵ�ֶ��ؽ�������(��Ǧ��ͭ��п����)��������[8]�����⣬����������ڵIJ���Ҳ���������绯����߰���ֲ������Ӫ������Ч��[2]�����磬�������ͨ�����������е�������Ӱ�������Ӻ������ӵ����䣬��ʹ�������ױ�ֲ���ϵ����[78-80]��
����(3)ϸ���ڽ���ø��ø�ٻ���
����ֲ��ϸ���ھ��е�����ԭ�����Ⱦ���澳������[81]����ֲ�ﲡԭ�����Ⱦ�Ͷ�ֳͨ���������ϸ���ڽ������[58]��ֲ�ﲡԭ����²������в���ϸ���ڽ���ø��(cellwalldegradingenzyme��CWDE)������ά��ø(cellulase)��ľ���ؽ���ø(lignindegradingenzyme)������ø(pectinase)�Ͷ�۰�����ȩ��ø(polygalacturonase��PG)�ȣ��������ֲ����ʲ��ϸ���ڣ�ʵ����Ⱦ����չ[82]�����Ų�ԭ������Ⱦ����ֳ������Ļ��۵���CWDE������ǿ�������ٽ�������뼰������չ[83]��
�������磬������ڵ�PG���������������Ͷ�ֳ����������ڵIJ����ܹ���������֯��pHֵ����PG��������ˮƽ[84-85]�����͵�����Ӫ����ֲ�ﲡԭ���(����̾��ͻ�ù��)ͨ�����ڲ���ɽ���Ⱦ��λϸ������pHֵ����5.0���ң������������������CWDE(��PG)��������pHֵһ��С��6.0[86]��������ᷢ��Эͬ���ã�ʹ����˿�˾�(.solani)������С�˾�(S.rolfsii)���ڵ�PG����ˮ�����ֲ������㣬�����ڲ�ԭ��������[87]��
�������⣬�������Ҳͨ������ֲ��ϸ���ڵĸ�����(Ca2+)�γɲ���ƾ��壬��������������������ϸ�����ʲ�Ļ��������ٻ�����֯��ϸ���ڽ��⣬�ƻ�����ֲ��ϸ���������Լ����߷�Ӧ[2,88]�����ỹ�ܹ�Ӱ��ֲ��������������PG���Եĵ���(��polygalacturonase-inhibitingprotein��PGIP)���ԣ��Ӷ��ӱ�ֲ�������Ӧ[89]��
����(4)�յ�����������ֲ�ﲡԭ�����Ⱦ���������У��������þ���������[5]��һ���棬��������Ⱦ�������յ�����ֲ��ROS�������ٽ����̾�����Ⱦ[66];��һ���棬�������Ƽ���ֲ����������Ӧ���ı��������������ԭˮƽ���������������Ӧ���յ�����PCDʵ���������Ⱦ����չ[68]�����磬���̾����ڵIJ��ή�ͻ���ϸ������pHֵ�����ϸ����������Ƽ�������������Ӧ����ֱ��������������صĶ������ø(polyphenoloxidase)���������ø�����ʶ���ź�ת�������ͼ����Ŀ���[21,90]��
�������⣬���̾���Ⱦ�����У����ᷢ�Ӷ�̬�������á����̾���Ⱦ���ڣ���������ROS������������(callose)�����ȷ�����Ӧ;����Ⱦ���ڣ������յ�������ROSˮƽ���ߣ��շ�����ϸ��PCD��Ӧ���ٽ������Բ�ԭ���ڼ����ڵĽ�һ����չ[63]����������ˮƽ����Ҳ����������ͨ������ROS��ֲ�D���ź����磬�յ�����ֲ���������[72]����ˣ���Ũ�Ȳ��������ڿ���ù���ķ���Ʒ��ѡ��[91]��
����5������ػ�����
����ֲ�ﲡԭ����IJ���ϳɼ�������ص��ػ���Ĺ����о���չ����ҪӰ������IJ����л�������ͷ������²��Եȡ����У��������ϳ���ػ����о����������̾�Ssoah����ͻ�䵼�²����������Ҿ����γ�����[68]�����ڣ����о�����microRNA-likeRNAsͨ�����������ػ���(��oah)������̾�����ϳ������[92]����ù��BcOah�����ó����²��Լ����Ͷ��Խ���[28]����ƿ����߾�(Cytosporachrysosperma)��CcOahȱʧ���������ϳɲ��ᡢ���������������١�������������[40]��������ή����ػ��������Ҫ�����ã�������̾�Ssodc2�����ó��������²�����������[93]��
�������⣬һЩ����ҲӰ���������IJ�����������̾�SsCaf1��SsSOD1�����ó��������ڵ����������½��������������²��Ե�Ҳ��������[94-95]�����̾���ɫ�غϳ�;��SCD1��THR1����ȱʧ���ܹ����Ͳ���IJ�����Ӱ����˷���[96]������ùOAH�����ó������ĺϳ���ȫ�����[27]��������oahA����ĺ���ù�IJ������ԼΪҰ���;����2��[33]��
����6չ��
����������ڵIJ�����ж��ֹ��ܺ����ã��������������������Ⱦ����ֲ��ȷ��������Ҫ���塣�������������������������ԡ���л;����������������ࡢ����Ӱ�켰��ػ����ܵ��о���չ���ܽ��˲���������������������²����Ƶȷ������Ҫ���ã���������������ϵͳ���о����д���ǿ��δ����������о��Ŀ�չӦ��ע���¼������棺
����(1)��������������Դ���������÷�Χ�д���չ���ر���ģʽ�������Ҫֲ�ﲡԭ���;(2)�������������ϳ�;�������ػ������д���ȷ�����������;��Դ��TCA��GAC��������δ������(3)����������������������д�ϵͳ�Կ�չ��������ͨ����ѧ���������������Ϣ�����������������Ҫ���ӣ����ö��ּ���(������ó���RNAi��Ĭ��CRISPR/Cas��)��ȷ�����ܵ�;(4)���������ֲ�ﲡԭ�����������ﲡ����ɫ���ط������ޣ�����ũҩ����ʹ�á�������Ⱦ����ҩ�Է��������ⲻ�ϳ��֣��ӿ�̽��ֲ�ﲡԭ������²���������Ҫ�²�����(�����)������ѧ���ܾ�����Ҫ���塣
����REFERENCES
����[1]NoonanSC,SavageGP.Oxalatecontentoffoodsanditseffectonhumans[J].AsiaPacificJournalofClinicalNutrition,1999,8(1):64-74
����[2]DuttonMV,EvansCS.Oxalateproductionbyfungi:itsroleinpathogenicityandecologyinthesoilenvironment[J].CanadianJournalofMicrobiology,1996,42(9):881-895
����[3]ConnollyJH,JellisonJ.Calciumtranslocation,calciumoxalateaccumulation,andhyphalsheathmorphologyinthewhite-rotfungusResiniciumbicolor[J].CanadianJournalofBotany,1995,73(6):927-936
����[4]LibertiD,RollinsJA,DobinsonKF.PeroxysomalcarnitineacetyltransferaseinfluenceshostcolonizationcapacityinSclerotiniasclerotiorum[J].MolecularPlantMicrobeInteractions,2013,26(7):768-780
����[5]PalmieriF,EstoppeyA,HouseGL,LohbergerA,BindschedlerS,ChainPSG,JunierP.Oxalicacid,amoleculeatthecrossroadsofbacterial-fungalinteractions[J].AdvancesinAppliedMicrobiology,2019,106:49-77
����[6]DerissenJL,SmithPH.Refinementofthecrystalstructuresofanhydrousα-andβ-oxalicacids[J].ActaCrystallographicaSectionBStructuralCrystallographyandCrystalChemistry,1974,30(9):2240-2242
�������ߣ�����껣�����������ӱ�������ɣ�����骣����������

ת����ע�����Է���ѧ����������http://www.fbxslw.com/wslw/28689.html